Project: Ecological Stoichiometry of Forest Food Webs
Overview
Ecological stoichiometry, is the study of how interactions between organisms and their environments are influenced by the balance of elements and energy within the environment (Sterner & Elser, 2002; Austin & Vitousek, 2012). Concepts and Principles of ecological stoichiometry allow us to quantify ecosystem processes such as nutrient cycling, resource selection, predation, herbivory, animal movement, and energy flow within food webs (Schade et al., 2005). This project takes an ecological stoichiometric approach to investigate a vegetation-snowshoe hare-lynx food web.
Geographically this study is location within the Central Newfoundland Forest Ecoregion. Sampling will occur within Terra Nova National Park (TNNP), which will serve as a reference landscape undisturbed by recent forestry activity, and within adjacent Forest Management Areas (FMAs) where commercial and domestic forest harvest is permitted.
Geographically this study is location within the Central Newfoundland Forest Ecoregion. Sampling will occur within Terra Nova National Park (TNNP), which will serve as a reference landscape undisturbed by recent forestry activity, and within adjacent Forest Management Areas (FMAs) where commercial and domestic forest harvest is permitted.
Graduate Students:
Landscape Applications of Ecological Stoichiometry in Boreal Forest EcosystemsTravis Heckford
MSc Candidate in the Landscape Ecology and Spatial Analysis Lab 2016 - Present The primary focus of my research project is to explore forest stoichiometry at multiple spatial extents (sample plot, forest patch, and forest stand) and to construct forest stoichiometric distribution models (FSDMs) (Figure 1) to identify important trophic interaction areas with regards to snowshoe hare and canada lynx. My metric of forest stoichiometry will be based on the elemental composition of important understory species foraged by snowshoe hare: blueberry (Vaccinium angustofilium), red maple (Acer rubrum), balsam fir (Abies balsamea), and white birch (Betula papyrifera). Using spatial models of forest stoichiometric distribution, and habitat suitability models for snowshoe hare and Canada lynx, we can identify areas of overlap and test for trophic interactions (Figure 2). There are different determinants of habitat selection at different scales. For instance, at the forest patch scale lynx may be driven by prey abundance, but at the forest stand scale, it may be driven by prey catchability (Figure 3). Using a multiple scale approach, I will test the prey abundance and catchability hypotheses. At the forest stand scale, I expect habitats with high forest stoichiometry will produce lower trophic interactions due to healthy hares being more likely to escape predation. If areas of low and high forest stoichiometry are identifiable I expect to observe high occurrences of trophic interactions in low stoichiometry areas and low occurrences in high stoichiometry areas. At the forest patch scale, I expect habitats with high forest stoichiometry will produce an abundance of hares and that lynx will spend more time in these areas hunting.
So What?Boreal forest ecosystems, although comprised of a heterogeneous matrix of forest patches, provide ideal conditions to test a large extent ecological stoichiometry study due to the fact that we can quantify and describe forest patches as discrete units. Industrial and land management conservation decisions are often informed by survey observations, species distribution models, and habitat suitability models. Using forest stoichiometric distribution models as a proxy of resource quality we can refine current species distribution models to realize habitat subtleties: trophic interaction areas.
Using a multi-scale integrated approach to construct forest stoichiometric distribution models allows us to address issues at difference levels of inference. These models will provide us with the opportunity to perceive the world with new boundaries. We will be able to identify edges and boundaries of nutrient flow, identify high and low spots of stoichiometry, address issues of landscape connectivity, and inform ecological research on herbivore behavior, population dynamics, and distribution. Results of this study can be used in boreal forest management situations across Canada. |
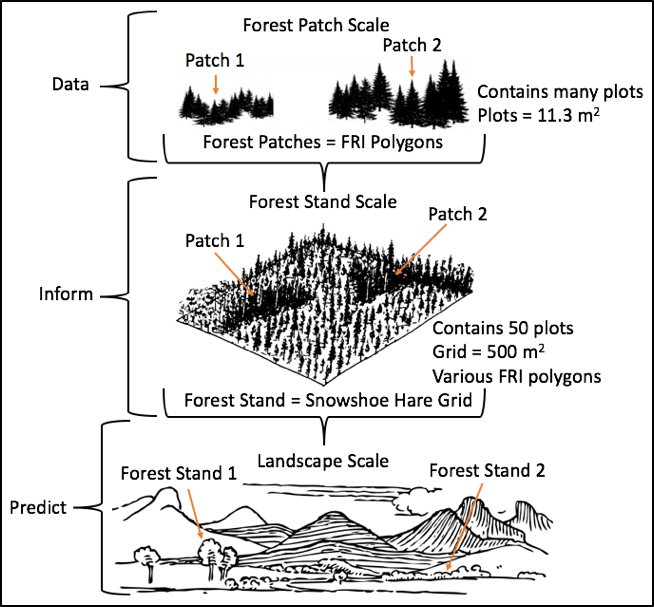
Figure 1. project design schema; data collected at the forest patch scale (FRI polygons) across many forest patches will be used to model forest patch stoichiometry at the landscape scale.
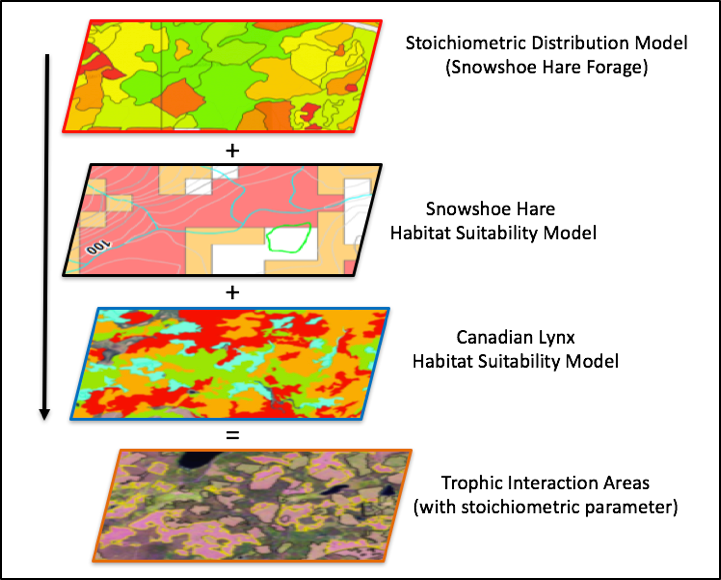
Figure 2. GIS approach to identifying areas of trophic interactions that include a stoichiometric distribution parameter.
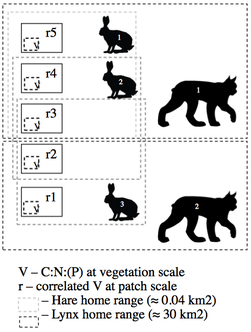
Figure 3. Lynx interaction schema based on different areas/regions of spatial stoichiometry.
|
Biochemical Dynamics of Food Webs
Matteo Rizzuto
PhD Student in the Ecosystem Ecology Lab
2016 - Present
PhD Student in the Ecosystem Ecology Lab
2016 - Present
|
My current research focuses on consumer-resource interactions and their effects on food web dynamics. I am using a plant-snowshoe hare study system to answer questions of intraspecific variability in body chemical composition and how this, in turn, affects landscape use and trophic web bioenergetics. To address these questions, I am using a multi-disciplinary approach that combines Ecological Stoichiometry — the study of how the nutrient content of organisms shapes their ecology — with Spatial Ecology and Wildlife Biology.
The first goal of my work is to describe how an individual’s development influences its nutritional requirements, by modifying its body chemical composition. This part of my work will focus on snowshoe hares (Lepus americanus), a fundamental herbivore in the boreal forest of North America. Numerous studies over the last two decades investigated individual variability in chemical composition, finding that it relates to body size (El-Sabaawi et al. 2012), diet (Gonzalez et al., 2011) and growth (Boros et al. 2015). So far, however, most of the research on this topic has been confined to aquatic species, or invertebrates. My results will shed new light on the ecology of this species, and also provide us with a framework to predict the chemical composition of individual hares (Fig. 4), which could be useful in future work focused on a hare’s predators - namely lynxes and coyotes. I will then combine this information with data on the availability and quality of resources to model individual space use decisions. The different quality of resources available over the landscape can generate hot- and cold-spots of nutrient (Carbon, Nitrogen, Phosphorus) abundance (Jean et al., 2015; Fig. 5). This, in turn, affect how consumer use their habitat (Moore et al., 2010; Nie et al., 2015). In general, herbivores tend to prefer areas where nutrient are more concentrated (Ball et al., 2000), and hence resource quality is higher, or when those chemical elements are more easily available (Youngentob et al., 2011). Using a combination of Forest Stoichiometric Distribution Models (see above), Resource Selection Function (Manly et al. 2002) and radio-tracking data from collared snowshoe hare, I will investigate the space use patterns of snowshoe hares (Fig. 6). So what?In addition to furthering our understanding of the dynamics of this environment and of the ecology of the species inhabiting it, results from this project will also be instrumental in informing a wide range of human activities — from logging practices to wildlife management and conservation policies. By providing an in-depth look into what drives foraging and habitat preferences of snowshoe hares, my work will improve our understanding of the ecology of this keystone species, as well as produce valuable knowledge that could inform future hunting, forestry and conservation practices.
|
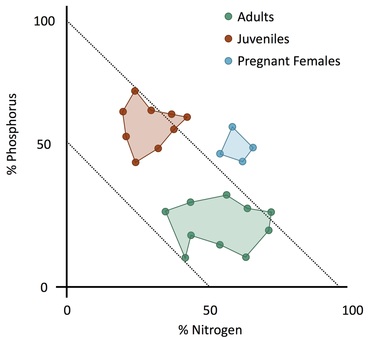
Fig. 4. Theoretical representation of the differences in body elemental content of three snowshoe hare age group. This figure uses the Right-Angle Mixture Triangle (Raubenheimer 2011) to express different whole-body concentrations of chemical elements relative to each other. The content of Carbon is shown in the implied Z axis, and is represented by the two dotted isoclines.
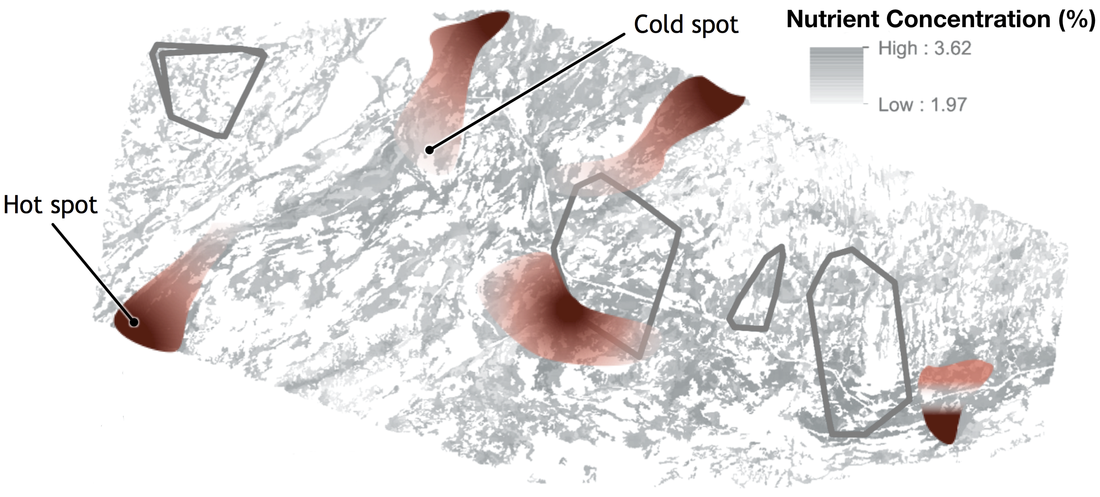
Fig. 5. Patterns of nutrient quality over the landscape. Hot-spots are areas where nutrient are highly concentrated, vice versa for cold-spots.
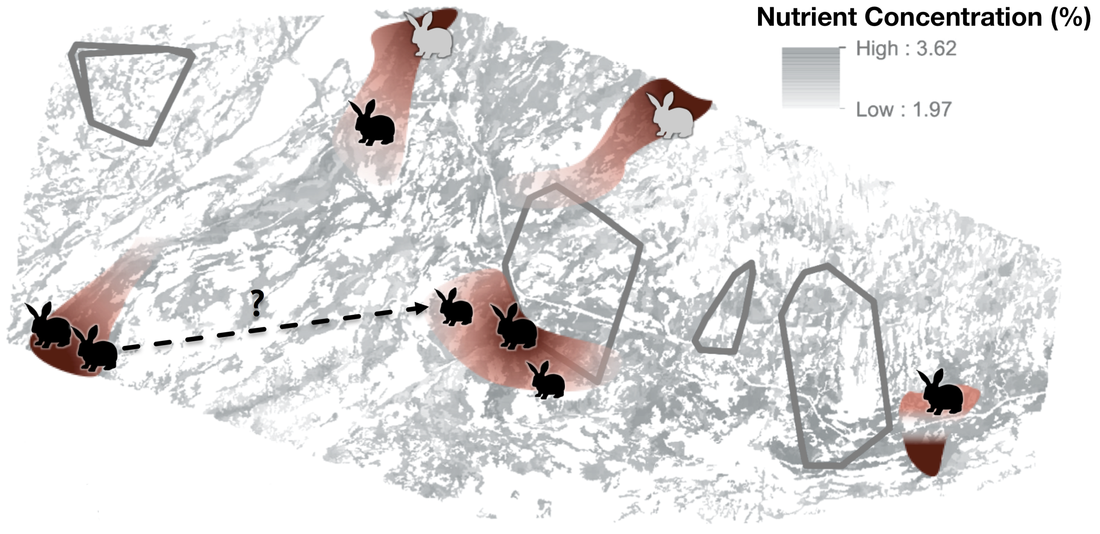
Fig. 6. Theoretical representation of how a pattern of nutrient hot- and cold-spots could affect snowshoe hare space use at the landscape level. Combining FSDM, RSF and radio tracking will provide strong theoretical and empirical basis to test this hypothesis.
|
References
Austin, A. T., & Vitousek, P. M. (2012). Introduction to a Virtual Special Issue on ecological stoichiometry and global change. New Phytologist, 196(3), 649–651.
Ball, J.P., Danell, K. & Sunesson, P. (2000). Response of a herbivore community to increased food quality and quantity: an experiment with nitrogen fertilizer in a boreal forest. J. Appl. Ecol., 37, 247–255
Boros, G., Sály, P. & Vanni, M.J. (2015). Ontogenetic variation in the body stoichiometry of two fish species. Oecologia, 179, 329–341
El-Sabaawi, R.W., Zandona, E., Kohler, T.J., Marshall, M.C., Moslemi, J.M., Travis, J., et al. (2012). Widespread intraspecific organismal stoichiometry among populations of the Trinidadian guppy. Funct. Ecol., 26, 666–676.
González, A.L., Fariña, J.M., Kay, A.D., Pinto, R., Marquet, P.A., Gonz??lez, A.L.A.L., et al. (2011). Exploring patterns and mechanisms of interspecific and intraspecific variation in body elemental composition of desert consumers. Oikos, 120, 1247–1255
Jean, P.-O., Bradley, R.L., Tremblay, J.-P. & Cote, S.D. (2015). Combining near infrared spectra of feces and geostatistics to generate forage nutritional quality maps across landscapes. Ecol. Appl., 25, 1630–1639
Manly BF, McDonald L, Thomas D, McDonald TL, Erickson WP. Resource selection by animals: statistical design and analysis for field studies. Springer Science & Business Media; 2007.
Moore, B.D., Lawler, I.R., Wallis, I.R., Beale, C.M. & Foley, W.J. (2010). Palatability mapping: a koala’s eye view of spatial variation in habitat quality. Ecology, 91, 3165–3176
Nie, Y., Zhang, Z., Raubenheimer, D., Elser, J.J., Wei, W. & Wei, F. (2015). Obligate herbivory in an ancestrally carnivorous lineage: The giant panda and bamboo from the perspective of nutritional geometry. Funct. Ecol., 29, 26–34
Schade, J. D., Espeleta, J. F., Klausmeier, C. A., McGroddy, M. E., Thomas, S. A., & Zhang, L. (2005). A conceptual framework for ecosystem stoichiometry: balancing resource supply and demand. Oikos, 109(1), 40–51. http://doi.org/10.1111/j.0030-1299.2005.14050.x
Sterner, R.W. & Elser, J.J. (2002). Ecological stoichiometry: the biology of elements from molecules to the biosphere. Princeton University Press
Youngentob, K.N., Wallis, I.R., Lindenmayer, D.B., Wood, J.T., Pope, M.L. & Foley, W.J. (2011). Foliage chemistry influences tree choice and landscape use of a gliding marsupial folivore. J. Chem. Ecol., 37, 71–84
Ball, J.P., Danell, K. & Sunesson, P. (2000). Response of a herbivore community to increased food quality and quantity: an experiment with nitrogen fertilizer in a boreal forest. J. Appl. Ecol., 37, 247–255
Boros, G., Sály, P. & Vanni, M.J. (2015). Ontogenetic variation in the body stoichiometry of two fish species. Oecologia, 179, 329–341
El-Sabaawi, R.W., Zandona, E., Kohler, T.J., Marshall, M.C., Moslemi, J.M., Travis, J., et al. (2012). Widespread intraspecific organismal stoichiometry among populations of the Trinidadian guppy. Funct. Ecol., 26, 666–676.
González, A.L., Fariña, J.M., Kay, A.D., Pinto, R., Marquet, P.A., Gonz??lez, A.L.A.L., et al. (2011). Exploring patterns and mechanisms of interspecific and intraspecific variation in body elemental composition of desert consumers. Oikos, 120, 1247–1255
Jean, P.-O., Bradley, R.L., Tremblay, J.-P. & Cote, S.D. (2015). Combining near infrared spectra of feces and geostatistics to generate forage nutritional quality maps across landscapes. Ecol. Appl., 25, 1630–1639
Manly BF, McDonald L, Thomas D, McDonald TL, Erickson WP. Resource selection by animals: statistical design and analysis for field studies. Springer Science & Business Media; 2007.
Moore, B.D., Lawler, I.R., Wallis, I.R., Beale, C.M. & Foley, W.J. (2010). Palatability mapping: a koala’s eye view of spatial variation in habitat quality. Ecology, 91, 3165–3176
Nie, Y., Zhang, Z., Raubenheimer, D., Elser, J.J., Wei, W. & Wei, F. (2015). Obligate herbivory in an ancestrally carnivorous lineage: The giant panda and bamboo from the perspective of nutritional geometry. Funct. Ecol., 29, 26–34
Schade, J. D., Espeleta, J. F., Klausmeier, C. A., McGroddy, M. E., Thomas, S. A., & Zhang, L. (2005). A conceptual framework for ecosystem stoichiometry: balancing resource supply and demand. Oikos, 109(1), 40–51. http://doi.org/10.1111/j.0030-1299.2005.14050.x
Sterner, R.W. & Elser, J.J. (2002). Ecological stoichiometry: the biology of elements from molecules to the biosphere. Princeton University Press
Youngentob, K.N., Wallis, I.R., Lindenmayer, D.B., Wood, J.T., Pope, M.L. & Foley, W.J. (2011). Foliage chemistry influences tree choice and landscape use of a gliding marsupial folivore. J. Chem. Ecol., 37, 71–84
